


Tat-NR2B9c
In the ever-evolving landscape of biomedical research, a new star is rising, capturing the attention of scientists and researchers worldwide. This star is none other than Tat-NR2B9c, a peptide whose unique properties and potential applications are setting the stage for groundbreaking advancements in medicine and therapeutic treatment.
At its core, Tat-NR2B9c is a synthetic peptide, meticulously designed to harness the power of cellular penetration and delivery mechanisms. Its origins can be traced back to the human immunodeficiency virus (HIV) Tat protein, renowned for its ability to cross cellular membranes with remarkable efficiency. By leveraging this intrinsic property, scientists have engineered Tat 9R2B9c to serve as a versatile tool in the delivery of therapeutic agents directly into cells, a feat that has long posed a challenge in the realm of drug development.

The Tat-NR2B9c peptide has shown clinical efficacy as a neuroprotective agent in acute stroke. Tat-NR2B9c is designed to prevent nitric oxide (NO) production by preventing postsynaptic density protein 95 (PSD-95) binding to N-methyl-D-aspartate (NMDA) receptors and neuronal nitric oxide synthase; however, PSD-95 is a scaffolding protein that also couples NMDA receptors to other downstream effects. Here, using neuronal cultures, we show that Tat-NR2B9c also prevents NMDA-induced activation of neuronal NADPH oxidase, thereby blocking superoxide production. Given that both superoxide and NO are required for excitotoxic injury, the neuroprotective effect of Tat-NR2B9c may alternatively be attributable to uncoupling neuronal NADPH oxidase from NMDA receptor activation.
To determine whether NR2B binding to PSD-95 is required for NMDAr-triggered superoxide production, we treated neuronal cultures with 100 μmol/L NMDA for 30 minutes together with Tat-NR2B9c over a range of concentrations and evaluated superoxide formation by the dihydroethidium method. At the highest concentration (0.5 μmol/L), Tat-NR2B9c suppressed NMDA-induced superoxide production by about 75% (Figures 1A and 1B). This effect was comparable to that achieved with the gp91ds-Tat peptide (Figures 1B and 1C), which prevents assembly of the active neuronal NOX2 complex.10,12 By contrast, the nNOS inhibitor L-NG-nitroarginine methyl ester (L-NAME) had no effect on superoxide production. The inhibitory effect of Tat-NR2B9c on NMDA-induced superoxide production was further confirmed by imaging formation of the lipid peroxidation product, 4-hydroxynonenal (Figures 1D and 1E).

Figure 1: Tat-NR2B9c blocks N-methyl-D-aspartate (NMDA)-type glutamate receptors (NMDAr)-triggered superoxide production.
Figure 1. Tat-NR2B9c blocks N-methyl-D-aspartate (NMDA)-type glutamate receptors (NMDAr)-triggered superoxide production. (A) Concentration-response curve. (B and C) Oxidized dihydroethidium (Eth fluorescence) in neuronal cultures. Scale bar = 20 μm. NMDA, 100 μmol/L; Tat-NR2B9c, 0.5 μmol/L; scrambled-sequence tat-conjugated peptide, 0.5 μmol/L; L-NG-nitroarginine methyl ester (L-NAME), 10μmol/L; gp91ds-Tat, 0.5 μmol/L. (D and E) Immunostaining (red) shows 4-hydroxynonenal formation. Green identifies neuronal processes (MAP-2), and in merged images, blue identifies cell nuclei (DAPI, blue). All studies with n > 3; *P < 0.05 versus NMDA.
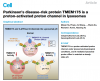
The dissociation of the NR2B–PSD95–nNOS signaling complex prevents the NMDAR-mediated production of the neurotoxic molecule NO, reducing stroke damage. (a) The NR2B subunit (orange) of NMDAR forms a multimeric protein complex with PSD95 and nNOS. (b) Tat-NR2B9c dissociates PSD95–nNOS from NR2BRs by disrupting the NR2B–PSD95 interaction. (c) ZL006 dissociates nNOS from NR2B–PSD95 by disrupting the PSD95–nNOS interaction as shown in following diagram:

As research into Tat 9R2B9c continues to unfold, its potential applications seem boundless. From combating challenging diseases like cancer and neurodegenerative disorders to enabling the development of targeted gene therapies, the possibilities are as vast as they are exciting. However, as with any emerging technology, the journey from the laboratory to clinical application is complex and fraught with challenges. Rigorous testing, validation, and refinement of Tat 9R2B9c-based therapies are essential steps on the path to realizing their full potential.
In order to observe the effects of Tat-9R2B9c in animals first, we used the PVO model that produces a small infarction (Figure 2A–B) without altering CT as compared with sham surgery animals (Figure 1Ai, iii). The animals were treated with vehicle (saline), control peptide Tat-NR2B9c(ADA), or low (0.3 nM/g) or high (3 nM/g) doses of Tat-NR2B9c(SDV) at 1 hour after PVO. Treatment with either vehicle or Tat-NR2B9c(ADA) resulted in cortical infarcts underlying the PVO, occupying approximately 9% to 10% of the hemisphere (Figure 2C). This was reduced by approximately 60% by Tat-NR2B9c(SDV) (3 nM/g; Figure 2C–D).

Figure 3: Effect of Tat-NR2B9c(SDV) posttreatment in the PVO model
Figure 3. Effect of Tat-NR2B9c(SDV) posttreatment in the PVO model. A, The 3 sites of PVO. B, Resulting typical infarct in triphenyltetrazolium chloride (TTC)-stained brain. C, Effect of the indicated drug and drug concentration on infarct size. N=8/group. Asterisk indicates significantly different from saline and Tat-NR2B9c(ADA) controls (ANOVA, P<0.05). D, Representative infarcts in TTC-stained coronal sections from each group. E–F, Effect of the indicated drug and gender on infarct size. SDV, Tat-NR2B9c(SDV); ADA, Tat-NR2B9c(ADA); n=8/group. Asterisk indicates different from saline control of the same gender (Student t test, P<0.01).
We also observe that Formalin induces central sensitization of deep dorsal horn WDR neurons is reduced by spinal pretreatment with Tat-NR2B9c. Formalin was injected into the hindpaw receptive field and the firing response of single WDR neurons was recorded (Figure 4c). This stimulus produces an afferent drive with a delayed central NMDA component to the response. In control recordings, where no drug was applied, formalin induced a characteristic biphasic neuronal firing response (Figure 4a; 1st phase: 0–10 minutes, total APs = 9,632 ± 1,678; 2nd phase: 10–70 minutes, total APs = 64,071 ± 16,220; n = 11). Control pretreatment with spinal Tat-NR2BAA (1.25 µg, n = 11), 20 minutes before formalin injection, did not alter this response (Figure 4a–c; 1st phase total APs = 13,970 ± 2,228, P > 0.05; 2nd phase total APs = 54,483 ± 13,673, P > 0.05). Spinal pretreatment with Tat-NR2B9c (12.5 ng, n = 10) did, however, significantly and selectively reduce 2nd phase neuronal firing (Figure 4a–c; total APs = 9,695 ± 5,386, P < 0.001), attributed to central sensitization of spinal dorsal horn neurons. Only a small nonsignificant reduction of neuronal firing during the 1st phase was observed following Tat-NR2B9c pretreatment, likely reflecting the inhibitory effects seen previously on postdischarge and wind-up. All cells recorded were characterized before injection of drug and formalin to ensure that cells were comparable between treatment groups.

Figure 4: Spinal Tat-NR2B9c reduces formalin-induced central sensitization.
Figure 4 Spinal Tat-NR2B9c reduces formalin-induced central sensitization. (a) Time course of firing of wide dynamic range (WDR) neurons following subcutaneous injection of formalin into the hindpaw receptive field with either no drug (control, n = 11) or following spinal pretreatment with Tat-NR2BAA (1.25 µg, n = 11) or Tat-NR2B9c (12.5 ng, n = 10, *P < 0.05 at 50 minutes versus Tat-NR2BAA). (b) Total neuronal activity during the 1st phase (0–10 minutes) and 2nd phase (10–70 minutes) of the response to formalin with no drug (control) or following spinal pretreatment with Tat-NR2AA or Tat-NR2B9c (2nd phase, ***P < 0.001, **P < 0.01 versus no drug and Tat-NR2BAA, respectively). (c) Example rate recordings of firing responses of WDR neurons to formalin with no drug (control) or following spinal pretreatment with Tat-NR2BAA (1.25 µg) or Tat-NR2B9c (12.5 ng). (d) Time course of pain-related behaviors following subcutaneous injection of formalin into the hindpaw following intrathecal pretreatment with Tat-NR2BAA (1.25 µg, n = 11) or Tat-NR2B9c (12.5 ng, n = 10, **P < 0.01 at 25 minutes versus Tat-NR2BAA; 125 ng, n = 6, ***P < 0.001 at 20, 25 minutes, *P < 0.05 at 30 minutes, **P < 0.01 at 35 minutes versus Tat-NR2BAA). (e) Total pain-related behavior during the 1st phase (0–10 minutes) and 2nd phase (10–60 minutes)of the response to formalin following spinal pretreatment with Tat-NR2AA or Tat-NR2B9c (2nd phase, 12.5 ng, **P < 0.01, 125 ng, ***P < 0.001 versus Tat-NR2BAA).
In conclusion, Tat-NR2B9c represents not just a scientific breakthrough, but a beacon of hope for the future of medicine. Its development underscores the importance of innovative research and the relentless pursuit of solutions to some of the most pressing medical challenges of our time. As we stand on the cusp of a new era in therapeutic delivery, the promise of Tat 9R2B9c serves as a testament to the power of science to change lives and shape the future.








Comments